曹林青,詹发强,杨 蓉,侯新强,包慧芳,侯 敏,王 宁,龙宣杞
(1.新疆大学生命科学与技术学院,乌鲁木齐 830046;
2.新疆农业科学院微生物应用研究所/新疆特殊环境微生物重点实验室,乌鲁木齐 830091)
【研究意义】新疆是我国最大的葡萄产区,2016年产量达267.6×104t左右,位居全国第一[1]。葡萄灰霉病(gray mold)是由灰葡萄孢(Botrytiscrnerea)引起的一种比较常见且危害严重的病害,由于葡萄采后的贮藏环境利于灰霉菌的生长,每年造成葡萄采后的腐烂损失率达30%以上[2]。国内外葡萄保鲜技术主要是低温结合二氧化硫(Sulfur Dioxide,SO2)防腐贮藏[3,4]。SO2的有效杀菌剂量和使葡萄受伤害的剂量相近,所以使用剂量不好控制,当SO2剂量过高时,葡萄抗氧化的酶促防御系统会受到损害,果实被漂白并产生异味[5]。嗜线虫致病杆菌(Xenorhabdussp.)是小卷蛾斯氏线虫(Steinernemacarpocapsae)的共生细菌,生存于侵染期线虫前肠的菌囊内。该菌随线虫的侵染进入寄主血腔,迅速大量增殖并产生多种代谢产物,为寄主线虫的生存提供合适的小生境。嗜线虫致病杆菌的这一生理代谢特性已引起重视,并已从代谢物中发现了几种新结构的抗生素[6]对细菌、酵母菌和真菌的生长具有广泛抑制活性[7, 8 ]。研究葡萄的采后贮藏保鲜技术,对提高鲜食型葡萄的贮藏保鲜有重要意义。【前人研究进展】培养的温度和曝气影响细菌的生长,影响抗生素的产生[9, 10]。Xenorhabdusspp为兼性厌氧菌,Chirayu Sa-uth等[11]通过响应面法优化了XenorhabdusstockiaePB09抗真菌活性的最佳培养基组成(蔗糖、酵母提取物、氯化钠和磷酸氢二钾等)。邱德文等[12]对致病杆菌D43菌株发酵培养基和发酵条件进行了研究,同时对该菌代谢过程pH值、还原糖、总糖、氨基氮与抗生素产量的关系进行了分析。接菌量较低时,菌体生长的迟滞期过长,致使培养发酵耗时较长。但接入过多体积的菌体时,可能会带进过多的代谢产物,产生的代谢产物反馈抑制,从而影响抑菌物质的产生[13]。在液体发酵培养的过程中,装液量和转速影响着菌株对氧气的需求程度[14]。【本研究切入点】现有葡萄灰霉病生防菌Xenorhabdusnematophila-ALL对其抑菌率相对不高且效价不稳定。需通过发酵条件优化以期提高其抑菌能力和水平。【拟解决的关键问题】对嗜线虫致病杆菌的液体发酵进行单因素和响应面法发酵优化,研究不同的培养基和不同的培养条件对灰葡萄孢的抑制作用,为开发和应用共生细菌Xenorhabdusnematophila-ALL生物防治葡萄灰霉菌奠定基础。
1.1 材 料
1.1.1 供试真菌
灰葡萄孢(Botrytiscinerea,简称B.cinerea)。
1.1.2 供试共生细菌
嗜线虫致病杆菌(ALL)保存于新疆农业科学院微生物应用研究所农用微生物实验室。
1.1.3 供试培养基
NBTA:营养琼脂45 g,溴百里酚兰 0.025 g,氯化三苯基四氮唑 0.04 g,蒸馏水1 L ;
NA培养基:牛肉膏3 g,蛋白胨5 g,琼脂18 g,蒸馏水1 L;
PDA培养基:马铃薯浸粉3.0 g,葡萄糖20.0 g,琼脂14.0 g,蒸馏水1 L。1×105Pa灭菌20 min;
TSY培养基:酵母膏5 g,TSB 40 g,蒸馏水1 L,pH7.2~7.4;
TSB培养基:大豆胨5 g,胰蛋白胨15 g,NaCl 5 g,蒸馏水1 L,pH7.2~7.4;
LB培养基:胰蛋白胨 0 g,酵母膏 5 g,NaCl 10 g,蒸馏水 1 L,pH7.2~7.4 ;
NB培养基:牛肉膏3 g,蛋白胨10 g,NaCl 5 g,蒸馏水1 L,pH7.2~7.4;
1.1.4 仪器与设备
MSSPX-250型生化培养箱,MLS-3020高压蒸汽灭菌锅,SW-CJ-2F型双人双面净化工作台,Eppendorf5418R离心机,OLYMPUS CX41相差显微镜,OLYMPUS SZ61体式显微镜,ZWY-2102C恒温培养振荡器。
1.2 方 法
1.2.1 共生菌分离
无菌操作台中,将由昆虫病原线虫致死的大蜡螟幼虫放入75%的乙醇中,浸没10 s左右,无菌滤纸吸去多余的酒精,剪开腹足,接种针沾取或挤出流出的液体至NBTA平板培养基上,28℃条件下暗培养36~48 h,直至出现能够吸附蓝色且周围有透明圈即为目标菌[15]。
1.2.2 无菌上清液制备
将培养好的蓝色Ⅰ型菌接种于NB培养基中,培养14 h得到种子液,以1%的比例接种于NB培养基中继续培养36 h,在4℃,10 000 r/min,离心10 min,将离心得到的上清液经0.22 μm滤膜过滤。
1.2.3 菌丝生长抑制率
将离心过滤处理过的无菌上清液,与已灭菌冷却至42℃的NA培养基以1∶100的比例混合均匀后倒入一次性培养皿中,每个处理4个重复。当对照组菌丝生长至培养皿边缘时测定各个处理的抑制率。
抑制率=(对照组菌块直径-试验组菌块直径)/(对照组菌块直径-菌饼直径)× 100%。
1.2.4 单因素试验
不同的培养基(NB、LB、TSB、TSY),不同的培养时间(24、48、72、96、120、144 h),种子液培养时间(12、16、20、24、28 h),接种量(1%,3%,5%,7%,9%),pH值(5,6,7,8,9),转速(160、180、200 r/min),装液量(10、20、30、40、50 mL/100mL)等因素对葡萄灰霉菌有显著抑制效果的因素为后续的响应面试验。
1.2.5 响应面优化设计
选取对试验影响较为显著的3个因素,利用Box-Behnken中心组合设计,进行三因素三水平响应面分析。
1.3 数据处理
采用Excel 2010计算数据,利用SPSS 22.0进行数据统计分析,使用Duncan多重比较检验法进行显著性分析。采用软件Design-Expert V8.0.5进行响应面的数据分析与处理。
2.1 单因素培养条件对ALL菌株发酵上清液抑菌活性的影响
2.1.1 培养基对发酵上清液抑菌活性的影响
研究表明,不同的培养基对共生细菌的影响较大,差异性显著(P<0.05)。TSY培养基对灰葡萄孢(B.cinerea)的菌丝抑制率最高,达48.12%。其次为TSB培养基,其抑制率为27.28%。LB培养基的抑制率最低,仅18.19%。TSB培养基与NB培养基的抑菌活性基本一致。因此将筛选出的TSY培养基作为后续对目标菌株ALL发酵条件优化的初始培养基。图1

图1 不同培养基发酵上清液下灰葡萄孢(B. cinerea)的抑制率变化
2.1.2 培养时间对发酵上清液抑菌活性的影响
研究表明,培养不同的时间对ALL菌株发酵上清液的抑菌活性差异性显著,24~120 h呈现出先上升后下降的趋势,且在72 h表现出最强的抑菌活性。24 h时的抑菌活性最低,呈现这种趋势的原因可能是在培养初期,摇瓶内菌体浓度暂时未达到最高值,随着时间的延长,菌体浓度升高,次生代谢产物量大幅积累,抑菌活性显著提高;
随着培养基内有限的营养和空间被用完,现抑制率降低。图2

图2 不同时间发酵上清液下灰葡萄孢(B.cinerea)的抑制率变化
2.1.3 种子液对发酵上清液抑菌活性的影响
研究表明,培养不同时间的种子对B.cinerea菌丝的抑制率不同,在12~28 h内,种子液的抑制率持续上升并在28 h抑制率达到最高,在12 h抑制率为0,16~24 h的种子液几乎无明显的抑制率,且在28 h的种子液抑制率显著高于其他时间的抑制率(P<0.05)。图3

图3 不同种子上清液下灰葡萄孢(B.cinerea)的抑制率变化
2.1.4 接菌量对发酵上清液抑菌活性的影响
研究表明,不同比例接菌量对灰葡萄孢(B.cinerea)菌丝生长无明显的显著差异性(P>0.05),5%的接菌量时,菌丝抑制率较高。图4

图4 不同接菌量下灰葡萄孢(B.cinerea)的抑制率变化
2.1.5 通气量对发酵上清液抑菌活性的影响
研究表明,各不同的装液量对灰葡萄孢(B.cinerea)菌丝生长的抑制率有显著性差异(P<0.05)。当装液量为50%时,对菌丝抑制率最高,达到51.8%,显著高于其他不同体积的装液量。图5
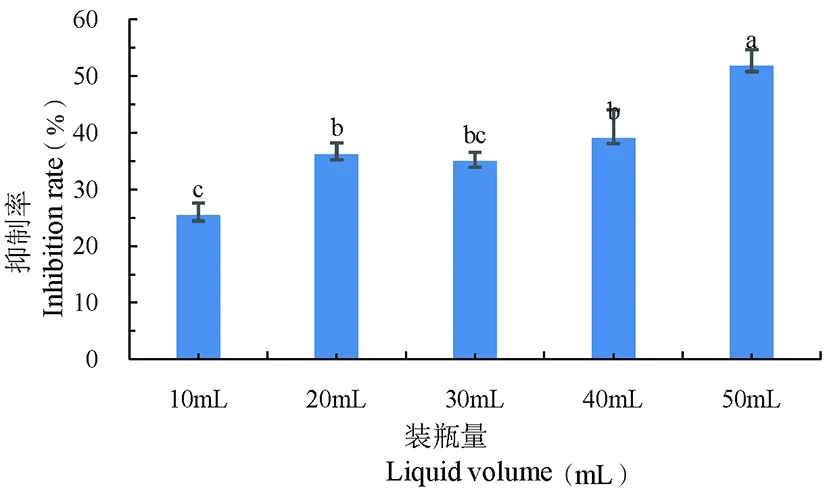
图5 装液量对灰葡萄孢(B.cinerea)抑制率变化
研究表明,转速在180 r/min时,对菌丝生长抑制率最高且显著高于其它两个转速的抑制率,达42.76%。图6

图6 不同转速下灰葡萄孢(B.cinerea)的抑制率变化
2.1.6 pH对发酵上清液抑菌活性的影响
研究表明,不同初始pH值对B.cinerea菌丝生长的抑制率不同,在初始pH为7时抑制率最高。起始 pH 直接影响共生细菌的生长环境,pH过高、过低都不利于菌体的生长。图7

图7 不同初始pH值下灰葡萄孢(B.cinerea)的抑制率变化
2.2 响应面因素优化分析
研究表明,拟合二次回归方程如下:
Y=+82.65+1.86A+4.59B+7.56C+0.052AB+2.47AC+0.28BC-21.41A2-5.84B2-6.74C2
Y是因变量(抑制率)
R2=0.981 2R2adj=0.957 0
F=40.53,P<0.01,差异极显著,模型建立有意义;
R2=0.981 2,回归方程拟合度良好,失拟项F=4.06,P>0.05,无显著性差异,非正常误差较小,得到灰霉菌的最佳抑制率。培养基初始pH值(A)、培养时间(B)、装液量(C)对灰霉菌抑制率的主次影响顺序:C>B>A。一次项B.C和二次项A2、B2、C2对响应值影响极显著,其余项对响应值影响不显著。表1~3

表1 因素水平编码

表2 Box-Behnken试验设计
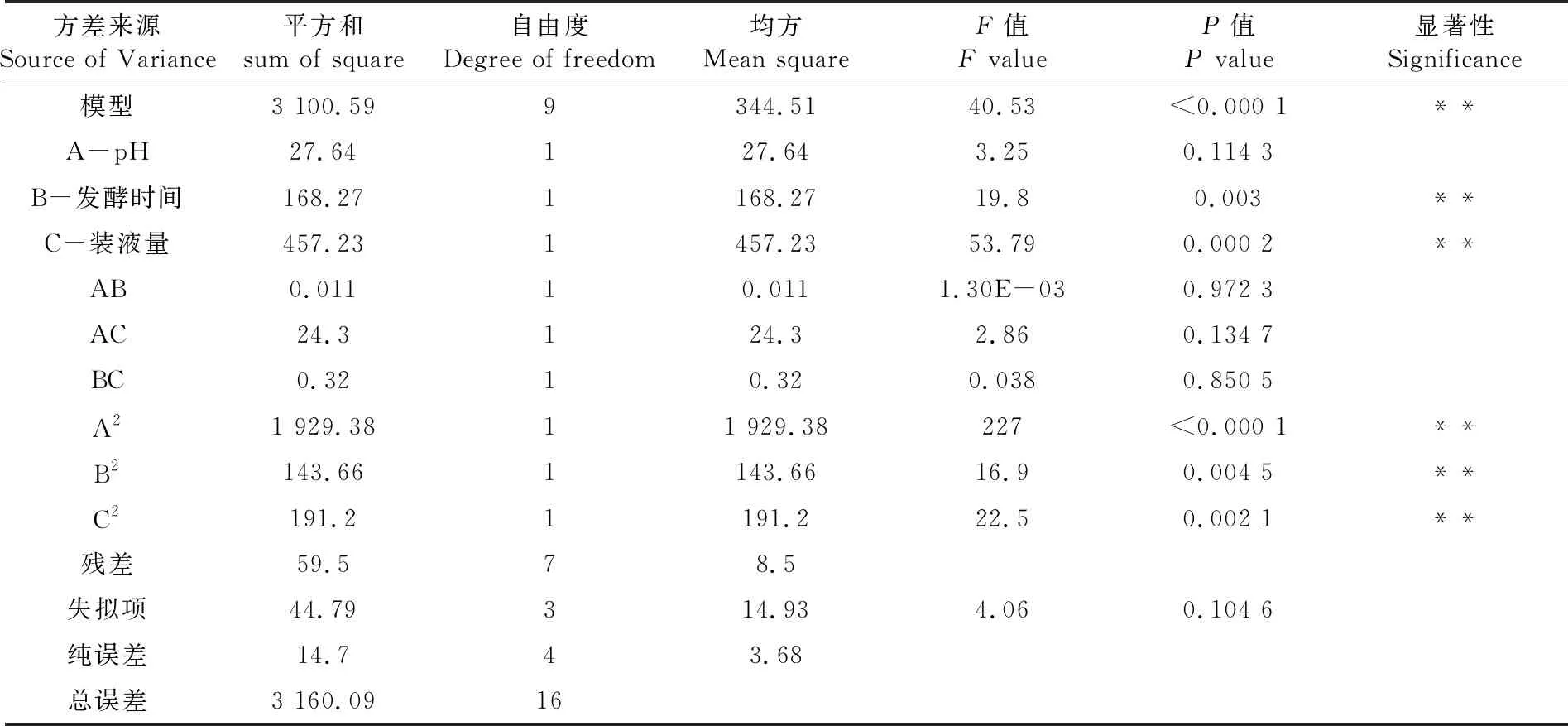
表3 回归模型方差
2.3 发酵时间与初始pH值对灰葡萄孢抑菌活性的影响
研究表明,当发酵时间恒定时,初始pH值在5~7时,抑制率呈现先增加后减少的趋势。同样地,当初始pH值一定时,随着发酵时间在60~84 h时,抑制率呈现先增加后减少的趋势。装液量水平为0时,发酵时间与pH交互作用的等高线侵向于圆形,二者交互作用无显著性。
当装液量恒定时,初始pH值在5~7时,抑制率呈现先增加后减少的趋势。当初始pH值一定时,装液量在40~60 mL/100 mL时,抑制率呈现先增加后减少的趋势。发酵时间水平为0时,装液量与pH交互作用的等高线更接近于圆形,装液量与初始pH值的交互作用不显著。
当装液量不变时,发酵时间在60 h与84 h之间时,抑制率呈现先增加后减少的趋势。当发酵时间一定时,在装液量在40与60 mL/100 mL时,抑制率呈现先增加后减少的趋势。初始pH值水平为0时,装液量与发酵时间交互作用的等高线非椭圆形,二者交互作用不显著。图8
在单因素优化试验中,Xenorhabdusnematophila-ALL在TSY培养基中发酵培养48 h后其抑制率最高,其次为TSB培养基,与马丽丽等[14]研究结果一致。要筛选出一种培养基作为发酵优化的初始培养基以期得到更多抑菌活性物质产量,4种培养基的配料成分分析,判断大豆胨,胰蛋白胨是产生抑菌物质的关键成分。通过对发酵时间的优化,拮抗菌对灰葡萄孢的抑制率呈现先上升后下降又上升的趋势,并在72 h时显示出最高的抑制率,产生这种现象的原因可能是与细菌利用营养物质有关,影响其抑菌物质的产生。接菌量会影响Xenorhabdusnematophila-ALL生长速度,摇瓶内细菌起始量越多的可能会较早达到最大值,能早合成细菌的代谢产物,但在研究中,不同的接菌量无显著差异。溶氧量影响发酵产物的产生,在液体摇床发酵培养过程中,装液量和转速影响着溶氧量,故两者需结合起来分析[16-19]。研究显示,当容积为50%时,转速为180 r/min时抑菌活性最高,表明了此条件为拮抗菌发酵的最佳通气量。pH作为共生细菌生长的重要环境因子,对其生长代谢及抑菌活性物质的产生有较大影响[20]。pH能影响细胞原生质的电荷,引起各种酶活性的变化,影响细菌底物的利用率和细胞结构,最终影响细胞的生长和产物的形成[21, 22]。研究发现,在培养基初始pH值为7.0时,抑制率最高,这与郭蔷薇等[23]在相同pH值条件下通过嗜线虫致病杆菌胞外产物对番茄灰霉病的结果一致。研究发现在中性环境中,Xenorhabdusnematophila发酵液能产生一些多肽,水溶性蛋白类抑菌物质,而在酸性和碱性环境中不利于该物质的产生。研究经过响应面分析结果发现目标菌株发酵上清液初始pH值,发酵时间,装液量等3个因素对灰葡萄孢(B.cinerea)菌丝生长的抑制率较高,通过响应面分析发现装液量是显著的影响因子,其次是发酵时间,而培养基初始pH值不显著。方香玲[24]对致病杆菌TB菌株进行了RSM发酵优化发现,筛选出最佳培养基为蛋白胨,培养条件为发酵时间(54.07 h),初始pH值(7.59)以及接菌量(9.95%)等3个因素,优化后提高了近73%,Wang等[25]对致病杆菌YL001菌株进行了RSM优化,确定了培养基最佳初始pH值7.64,装液量10%,转速220 r/min,接种量15%等,可达到最大的抗生素活性。

图8 各因素交互作用下B. cinerea 抑制率响应面图及等高线
最适培养基为TSY,最适培养时间为72 h,最适种子液为28 h,最适接菌量为5%,最适通气量为50 mL/100 mL、180 r/min,最适pH值为7.00。该优化条件为:培养基初始pH值为7.12,装液量56%,发酵培养时间为77 h,经验证抑菌率为87.70%,比优化前提高了1.82倍。
参考文献(References)
[1] 中国统计年鉴[J].北京:中国统计出版社,2017.
National Bureau of Statistics.ChinaStatisticalYearbook[J]. Beijing: China Statistics Press, 2017.
[2] 罗琳, 王其慧, 赵海霞, 等.葡萄灰霉病生防菌株的筛选及其拮抗菌机理初探[J].中国酿造, 2017, 36(4): 93-98
LUO Lin, L, Wang Qihui, ZHAO Haixia, et al. Screening of bio-control strain againstBotrytiscinereaand preliminary research on its antagonistic mechanisms [J].ChinaBrewing, 2017, 36(4): 93-98.
[3] Ding Y L, Yang X Z, Li X H, et al. Effect of SO2 Interval Fumigation on Storage Quality of Red Grape[J].AppliedMechanicsandMaterials, 2013, (411):3170-3173.
[4] 周拥军,陈文煊,陈杭君,等.SO2控释保鲜剂对南方巨峰葡萄的贮藏效果研究[J].食品科学,2008,29(7):445-447.
ZHOU Yongjun, CHEN Wenxuan, CHEN Hangjun, et al. Study on storage effect of SO2controlled release preservative on southern Jufeng grape[J].FoodScience, 2008, 29(7):445-447.
[5] Gabler F M, Smilanick J L, Mansour M F, et al.Influence of fumigationwith high concentrations of ozone gas on postharvest gray mold and fungicide residues on table grapes[J].PostharvestBiologyandTechnology, 2010, 55(2):85-90.
[6] McInerney B V, Greg son R P, Akhurst R J. Biologically active metabolites fromXenorhabdussp. [J].JournalofNaturalProduct, 1991, 54(3): 774-795.
[7]Akhurst R J. Antibiotic activity of Xenorhabdus spp. bacteria symbiotically associated with insect pathogenic nematodes of the families Heterorhabditidae and Steinernematidae [J].JournalofGeneralMicrobiology, 1982, (128): 3061-3065.
[8]Paul V J, Frautschy S, Fenical W, et al. Antibiotics in microbial ecology : isolation and structure assignment of several new antibacterial compounds from the insect - symbiotic bacteria Xenorhabdus sp. [J].JournalofChemicalEcology, 1981, (7): 589-597.
[9]Maxwell P, Chen G, Webster J M, et al. Culture conditions forXenorhabdusandPhotorhabdussymbionts of entomopathogenic nematodes [J].Nematologica, 1996, 42(1): 124-130.
[10]Fang X L, Han L R, Cao X Q, et al. Statistical Optimization of Process Variables for Antibiotic Activity ofXenorhabdusbovienii[J].PlosOne, 2012, 7.
[11] Sa-Uth C, Rattanasena P, Chandrapatya A, et al. Modification of Medium Composition for Enhancing the Production of Antifungal Activity from Xenorhabdus stockiae PB09 by Using Response Surface Methodology[J].InternationalJournalofMicrobiology, 2018:1-10.
[12] 邱德文,焦宁宁,刘峥,等.致病杆菌d43菌株产素培养基及发酵条件[J].中国生物防治,2006,(1):58-62.
QIU Dewen,JIAO Ningning, LIU Zheng, et al. Cultural Medium and Fermentation Conditions ofXenorhabdussp. Strain D43 [J].ChineseJournalofBiologicalControl, 2006,(1):58-62.
[13]刘洪敏, 钱海涛, 董辉, 等. 昆虫病原线虫共生菌5-5B培养特性及杀虫活性研究 [J]. 中国植保导刊, 2007, 27(4): 5-9.
LIU Hongmin, QIAN Haitao, DONG Hui, et al. Studies on the cultivation characteristics and insecticidal activity of anentomopathogenic nematode symbiotic bacteria strain 5-5B [J].ChinaPlantProtection, 2007, 27(4): 5-9.
[14] 马丽丽, 台莲梅, 许艳丽, 等. 发光杆菌NJ菌株的抑菌活性研究 [J]. 植物保护, 2008,34(3): 25-29.
MA Lili, TAI Lianmei, XU Yanli, et al. A study on the antifungal activity of Photorhabdus sp. NJ [J].PlantProtection, 2008, 34(3): 25-29.
[15] 詹发强, 布卡·欧尔娜, 侯敏, 等. 新疆嗜菌异小杆线虫及其共生细菌的初步鉴定研究[J]. 新疆农业科学, 2015, 52(2): 244-251.
ZHAN Faqiang, Buka Ouerna, HOU Min, et al. Preliminary Identification of Entomopathogenic Nematodes (Heterorhabditisbacteriophora strain Nileke) and Its Symbiotic Bacteria in Xinjiang [J].XinjiangAgriculturalSciences, 2015, 52(2): 244-251.
[16]Akhurst R J, Boemare N E. A numerical taxonomic study of the genusXenorhabdus(Enterobacteriaceae) and proposed elevation of the subspecies ofX.nematophilusto species [J].JournalofGeneralMicrobiology, 1988, 134(7): 1835-1845.
[17]Liu J, Berry R, Poinar G, et al. Phylogeny ofPhotorhabdusandXenorhabdusSpecies and Strains as Determined by Comparison of Partial 16S rRNA Gene Sequences [J].InternationalJournalofSystematicBacteriology, 1997, 47(4): 948.
[18]Boemare N E, Akllurst R J. Biochemical and physiological characterization of colony form variants inXenorhabdusspp. (Enterobacteriaceae) [J].JournalofGeneralMicrobiology, 1988, (134):751-761.
[19] Chen G H, Maxwell P, Dunphy G B, et al. Culture conditions for Xenorhabdus and Photorhabdus symbionts of entomopathogenic nematodes [J].Nematologica, 1996, (42):124-127.
[20] 郭树奇.pH调控嗜线虫致病杆菌抑菌活性机理的初步研究[D].杨凌:西北农林科技大学,2015.
GUO Shuqi.StudyonantibacterialmechanismofXenorhabdusnematophilaregulatedbypH[D].Yangling: Northwest A & F University, 2015.
[21] 周启, 王道本. 农用抗生素和微生物杀虫剂 [M]. 北京:中国农业出版社, 1995.
ZHOU Qi, WANG Daoben.Agriculturalantibioticsandmicrobialinsecticides[M]. Beijing: China Agriculture Press, 1995.
[22] 陈坚, 李寅. 发酵过程优化原理与实践 [M]. 化学工业出版社, 2002.
CHEN Jian, LI Yin.Theprincipleandpracticeoffermentationprocessoptimization[M].Beijing: Chemical Industry Press, 2002.
[23] 郭蔷薇,张淑静,李忝珍,等.恒定pH对XenorhabdusnematophilaYL001抑菌活性及抑菌物质合成相关基因表达的影响[J].西北农业学报,2016,25(9):1413-1419.
GUO Qiangwei, ZHANG Shujing, LI Tianzhen, et al. Effect of Constant pH on Antimicrobial Activity and Related Gene Expression of Antimicrobial Bioactivities Synthesis ofXenorhabdusnematophilaYL001 [J].ActaAgriculturaeBoreali-occidentalisSinica, 2016, 25(9):1413-1419.
[24] 方香玲. 昆虫病原线虫共生菌的鉴定、培养及其抑菌活性研究[D]. 杨凌:西北农林科技大学, 2008.
FANG Xianling.Studyonidentification,cultureandantimicrobialactivityofentomopathognicbacteria[D]. Yangling: Northwest A & F University, 2008.
[25] Wang Y H, Feng J T, Zhang Q, et al. Optimization of fermentation condition for antibiotic production byXenorhabdusnematophilawith response surface methodology [J].JournalofAppliedMicrobiology, 2010, 104(3):735-744.
猜你喜欢抑制率线虫菌株夏季蔬菜换茬期线虫防治要注意今日农业(2022年14期)2022-09-15植物根际促生菌Bacillus mycoides Gnyt1菌株生物学特性比较研究草地学报(2022年3期)2022-03-28双酶水解鱼鳞蛋白制备ACE抑制肽的工艺优化研究现代农村科技(2022年1期)2022-01-21菌株出马让畜禽污染物变废为宝今日农业(2021年11期)2021-11-27血栓弹力图评估PCI后氯吡格雷不敏感患者抗血小板药物的疗效中国循证心血管医学杂志(2021年10期)2021-11-05蔬菜使用不同处理方式对农药残留影响的研究食品界(2021年7期)2021-07-19毛木耳优良杂交菌株的选育*中国食用菌(2020年11期)2021-01-18基于培养池阵列微流控芯片的单线虫并行分离条件考察西北农业学报(2020年12期)2020-12-14我国松树体内线虫种类及其分布中国森林病虫(2019年5期)2019-10-11朝鲜孢囊线虫——浙江省孢囊线虫新记录种浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17